The XV Collection: Living l’AVIDA loca — origins of multicellularity

by Harmit Malik
Even before John Maynard Smith formalized the term, evolutionary biologists have been fascinated by major evolutionary transitions, including the transition from single-celled to multicellular organisms. Multicellularity occurs over and over in the evolutionary record, giving rise to the bounty of life-forms visible to the naked eye. Simple, undifferentiated multicellularity can be easily explained as an example of practical cooperation, wherein different single-celled entities pool resources in a multicellular commune while preserving their individuality and the right to procreate.
However, differentiated multicellularity involves a Faustian bargain, akin to eusocial insect societies with few queens and many workers. What gives rise to such reproductive division of labor between differentiated non-reproductive cells that make up the soma and the few reproductive germ cells that are capable of giving rise to new offspring? The ‘dirty work’ hypothesis proposed that the pressures of doing ‘work’ (cell division, metabolic activity) are inherently mutagenic, and therefore likely to compromise genomic integrity. Thus, sequestration of cells dedicated to the task of reproduction absolves them of ‘dirty work’ which keeps their genomes as pristine as possible, and thus leads to the germ-soma dichotomy.
Despite being elegant and powerful in its simplicity, the ‘dirty work’ hypothesis is nevertheless tricky to test since it would require capturing the transition to differentiated multicellularity in the act. In recent years, experimental evolution models have made significant advances in our understanding of transitions to undifferentiated multicellularity in many unicellular organisms (yeast, Volvox, Chlamydomonas, choanoflagellates) but reproductive differentiation remains elusive. Enter AVIDA, a powerful artificial life software platform designed by Charles Ofria, Chris Adami, and C. Titus Brown, to study the evolutionary biology of self-replicating and evolving digital cells, self-replicating computer programs that evolve in an open-ended fashion.
In a fascinating paper published in 2014 in PLOS Biology, Heather Goldsby and colleagues created a world of 400 multicells in the AVIDA platform. Each multicell is akin to a multicellular organism, and comprises individual digital cells, performing a combination of logic functions, which ultimately determines how many resources flow into the multicell, and thereby its ability to replicate. Into this system of multicellular ‘organisms,’ the authors introduced a mutation rate into individual cells, which allowed them to acquire new functions and therefore more resources but also increased the probability of ‘genomic’ damage. Intriguingly, as the authors vary the mutation rate, they find ‘somatic’ cells incapable of self-replication at intermediate mutation rates. Closer examination confirms that these somatic cells are indeed performing more metabolic functions (‘dirty work’) and therefore taking a bigger hit to their genome integrity, whereas ‘germ’ cells maintain their own, and therefore the multicell’s, genetic information in pristine condition.
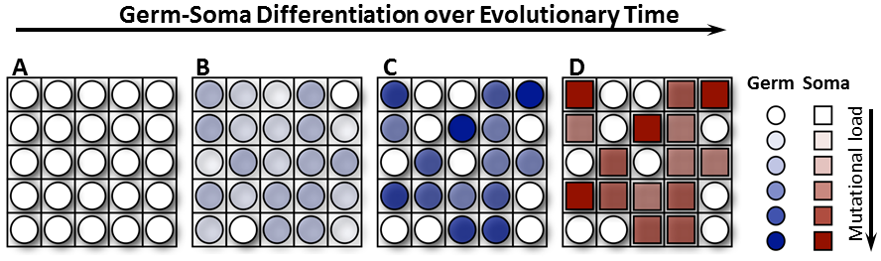
The authors exploit the power of AVIDA by following individual lineages of multicells (and their constituent cells) to find that multicells almost always first favor the development of ‘pseudo-somatic’ cells that perform much higher metabolic functions and therefore increase organismal (multicell) fitness. These ‘pseudo-somatic’ cells subsequently lose their ability to self-replicate, thereby avoiding a catastrophe of genomic mutation in the organism. Thus, a division of labor between ‘dirty work’ * and reproduction is the most successful long-term strategy for multicellular organisms. Indeed, acquisition of somatic cells also allowed multicells to explore previously forbidden phenotypic niches under conditions in which mutational load increases with every new function. One can easily imagine that an increase in body size and complexity may represent such a mutational ramp-up. Another elegant paper in PLOS Biology (Radzvilavicius et al., 2016) nicely demonstrates that preservation of mitochondrial genome integrity and quality is one example of how germ cells in a multicellular organism may avoid the cost of ‘dirty work’ paid by the soma.
Despite involving digital life-forms, experiments such as those performed by Goldsby et al. are an excellent example of the extraordinary insight they provide into evolutionary transitions that might otherwise be impossible to study with living, biological organisms. Co-operative systems such as multicellular organisms always operate under peril of cheaters, mutation-laden somatic cells that do not cede the reproductive role to pristine germ cells. However, the authors show that multicellular organisms comprising these high-mutation cells are less fit. Thus, their experiments not only provide valuable insight into ancient evolutionary transitions to multicellularity but may also guide studies of reversion to unicellularity, whereby cancerous cells arise by flouting rules governing replication or quiescence in multicellular organisms.
Goldsby HJ, Knoester DB, Ofria C, Kerr B (2014) The Evolutionary Origin of Somatic Cells under the Dirty Work Hypothesis. PLOS Biology 12(5): e1001858.https://doi.org/10.1371/journal.pbio.1001858
Radzvilavicius AL, Hadjivasiliou Z, Pomiankowski A, Lane N (2016) Selection for Mitochondrial Quality Drives Evolution of the Germline. PLOS Biology 14(12): e2000410.https://doi.org/10.1371/journal.pbio.2000410
Chase JM (2014) A Fool to Do Your Dirty Work?. PLOS Biology 12(5): e1001859. https://doi.org/10.1371/journal.pbio.1001859
Harmit Malik is the Principal Investigator at the Malik Lab at the Fred Hutchinson Cancer Research Center, and is a member of the PLOS Biology Editorial Board.

This blog post is the ninth in a series of twelve, forming PLOS Biology’s XV Collection, celebrating 15 years of outstanding open science; read Lauren Richardson’s blog for more information.
Featured image credit: pbio.2000410
Harmit Malik image credit: Aida de la Cruz